The old man’s beard leaf miner (Phytomyza vitalbae) was introduced into New Zealand in 1996 to help control the introduced clematis vine, old man’s beard (Clematis vitalba). However, old man’s beard leaf miners are parasitised in New Zealand, reducing their impacts on old man’s beard. There are seven known native and exotic parasitoids that parasitise the old man’s beard leaf miner and two of these also parasitise the closely related native clematis leaf miner (Phytomyza clematadi), which utilises several New Zealand native Clematis species. The diagram of this food web shows who eats whom.
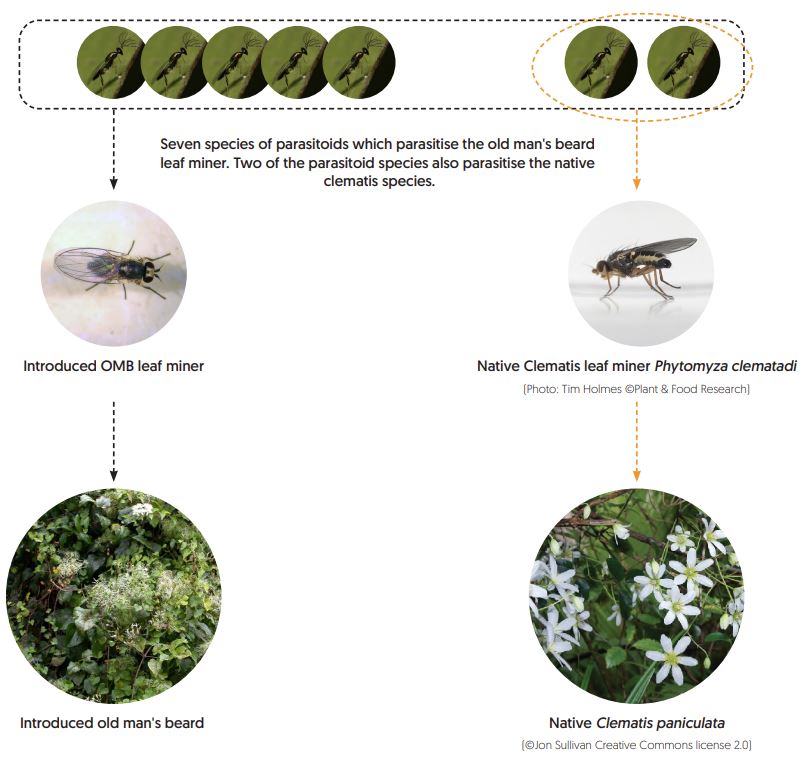
As part of a study investigating the non-target impacts of introduced weed biocontrol agents, researchers at MWLR have been working to untangle the impacts of the parasitoids and the introduced leaf miner on the native clematis leaf miner. Specifically, we investigated whether the introduction of the old man’s beard leaf miner has increased parasitism on the native leaf miner by increasing the abundance of their shared parasitoids (known as ‘apparent competition’).
Non-target impacts are impacts that do not relate to the target host plant, in this case old man’s beard. Non-target impacts can be direct, for example, if old man’s beard leaf miners feed on other plants instead of old man’s beard; or indirect, for example, through apparent competition if the presence of old man’s beard leaf miner has impacts on other food web links through intermediary species.
As an example of direct impacts, the old man’s beard leaf miner adults occasionally lay eggs (and larval mines can develop) on native Clematis species when old man’s beard and native clematis vines are growing close together (although the native Clematis species are a less preferred host). In a previous study investigating this non-target impact, we found no native leaf miners present on old man’s beard, and very few old man’s beard leaf miners on native Clematis species compared to old man’s beard. We also found that the introduced old man’s beard leaf miner is much more abundant on old man’s beard than the native leaf miner is on native Clematis species.
In the current study we investigated whether the introduction of the old man’s beard leaf miner is having indirect non-target impacts on the native clematis leaf miner by increasing the number of shared parasitoids in the environment, resulting in increased parasitism of the native leaf miner. To do this we checked vines of native Clematis species and old man’s beard for leaf miners and recorded the percentage of leaves mined. We then tried to determine the fate of each leaf miner to see whether they successfully developed into adult leaf miners (flies) or if they were parasitised. A total of 15,773 leaves on 81 vines (71 native and 10 old man’s beard) were checked, and 753 (294 native and 459 old man’s beard) mined leaves collected. Of the vines that were assessed, 5% of native Clematis species and 25% of old man’s beard leaves were mined.
Native clematis vines were selected at sites with and without old man’s beard present. At the sites where old man’s beard was not growing with native clematis vines, we measured the distance to the nearest old man’s beard. It was important to measure this for two reasons. Firstly, increased numbers of parasitoids may only occur close to old man’s beard, where the old man’s beard miner is often common. Secondly, we know from work by Quentin Paynter and colleagues that adult female old man’s beard leaf miner flies can only lay eggs if they have fed on the leaves of old man’s beard: feeding tests showed that the flies never feed directly on native clematis leaves. As a result, old man’s beard mines are unlikely to be found on native clematis unless the plants are close to old man’s beard plants.
We found old man’s beard leaf miner attacking only 0.14% of native clematis leaves, confirming previous conclusions that direct non-target impacts are rare. As with the previous study, we found a decline in old man’s beard leaf miner attack on native clematis as we moved further away from old man’s beard plants: beyond 1 km even the minor non-target attack we found vanished.
However, our main goal was to determine the levels of parasitoid attack in both the native and introduced leaf miners. To do this, we either diagnosed exit holes if leaf mines were vacant, or reared resident leaf miners through to adulthood. The larva of the old man’s beard leaf miner exits its mine and drops to the soil to pupate, leaving a tell-tale semi-circular flap behind at the end of its mine. In contrast, the larva of the native leaf miner pupates inside the end of the mine: the adult fly then emerges directly from the leaf mine, leaving a messy hole, and sometimes an empty pupal case behind. With both miner species, if the larva has been parasitised, the emerging adult parasitoid leaves a small round hole, which can be seen towards the end of the mine.
The big question for our study was whether the old man’s beard leaf miner could be harming the native leaf miner through apparent competition via the shared parasitoid species. Our hypothesis was that parasitoid numbers would be increased around old man’s beard plants because of the high levels of old man’s beard leaf miner, and that these parasitoids could then cause increased levels of attack on the relatively rare native clematis leaf miner – a potentially harmful indirect non-target impact on a native species.
The data suggest this is not happening. They show that parasitism of the native leaf miner did not increase when native Clematis species were growing close to old man’s beard. In fact, the native leaf miner appeared less likely to be parasitised when surrounded by an abundance of the old man’s beard leaf miner.
This result was unexpected. A possible explanation is that high densities of the old man’s beard leaf miner act as a local ‘sink’ for parasitoids rather than a source. Optimal foraging theory, for example, predicts parasitoids should concentrate searches in dense patches of prey, which are most likely to be old man’s beard leaf miner. To increase their success rates at finding prey, parasitoids may therefore avoid low density patches of leaf mines, meaning that the rarer native miners on native clematis tend to escape attack. This effect might be exacerbated because parasitoid success in attacking miners on old man’s beard may be higher (per unit of parasitoid search time) as it probably takes less time for the female wasp to drill into mines in the thinner, deciduous leaves of old man’s beard compared to the thicker, evergreen leaves of native clematis species.
Another fascinating contributing mechanism could be that relatively highly mined old man’s beard vines are producing more herbivore-induced volatiles (HIVs), the plant ‘cry for help’ hypothesis: the plant’s intention is that parasitoids will be attracted to these HIVs, resulting in higher parasitism levels where miners are abundant. There is a lot more research to undertake on biocontrol effects in this food web!
Regardless of the mechanism, we have shown that there are no negative food web effects on our relatively rare native clematis leaf miner, and perhaps even a small benefit. This is an important study because it demonstrates that food web impacts from introduced biocontrol agents should not be assumed to be always negative.
Further reading:
Paynter Q, Martin N, Berry J, Hona S, Peterson P, Gourlay AH, Wilson-Davey J, Smith L, Winks C, Fowler SV 2008. Non-target impacts of Phytomyza vitalbae a biological control agent of the European weed Clematis vitalba in New Zealand. Biological Control 44(2): 248–258.
This project is part of MWLR’s Beating Weeds Programme, funded by the Ministry of Business, Innovation and Employment’s Strategic Science Investment Fund in collaboration with B3 partners. Ruth Fleeson (Horizons Regional Council) is thanked for her help locating clematis vines.
Key contact
